Mammalian Toxicology, Session 13
Adequacy of models for developmental
toxicity; C 10, 29; Steroid disruptors: estrogen, androgen, progestin,
corticoid; C 20, 21, 29; Thyroid, retinoid and other disruptors; C 10,
20, 21; Take Home Exam Questions Distributed
Adequacy
of
models for developmental toxicity
The best
existing models for
developmental toxicity use multiple generations to evaluate impacts on
both the first and second generations following dosage of the parental
generation (usually of either the male or female parent only so as to
optimize the identification of the target of the actual insult).
Evaluations of the first generation indicate
direct effects of toxic insult on development. This may involve
germline
mutations in the parental generation if exposure took place in the
parents
prior to mating. It may involve genotoxic impacts involving the
germline or
embryonic F1 tissues. It may involve teratogenic
insults derived from
either
mutational or epigenetic influences within the F1
embryo. More subtle
impacts
on the F1 generation may appear later in development or
during aging,
e.g.,
increased incidences of particular tumor types. Evaluations of
the F2
generation
primarily serve to demonstrate the integrity of the F1
generation's
germinal
and reproductive tract. The model organisms used might include
common
test
species, but they may also include more rapidly reproducing animals
such
as zebra fish, a model for exploration of development within the
laboratory. Explorations may also include evaluation of mRNA
transcription or protein synthesis within specific tissues or cells
within the developing test organism.
Note however
that disruptions
of many
of the epigenetic influences on development (hormonal, nutritional) are
not
ascertained, or are observed only indirectly, in these tests.
Tests
that
look at shorter test segments or isolated tissues are further blinded
to
the complexities of the developmental process which is actually a three
body
problem (father, mother, offspring). As environmental exposures
of
populations become more important with respect to developmental insults
(see anti-estrogens and anti-androgens below) it will probably become
important to include test designs that explore a variety of non-protein
as well as protein hormones during the course of gestation and
development. Moreover, it will also be important to explore the
impacts
of exposure of both parents on the developmental outcomes of the
offspring.
Endocrine
Disruption
As a topic of
active interest
currently and in the recent past there are many references that can be
pointed to:
Silent
Spring. R.
Carson. Fawcett Crest: New York, NY. 1970. [Many other editions are
available.]
Our
Stolen Future. T.
Colborn, D. Dumanoski, J.P. Myers. Penguin Books: New York, NY. 1997.
Out Stolen Future - an
activist website
Hormonally
Active Agents in
the Environment. Committee on Hormonally Active Agents in the
Environment, Board on Environmental Studies and Toxicology, Commission
on Life Sciences, National Research Council. National Academy Press:
Washington, D.C. 1999.
Endocrine
and Hormonal
Toxicology. P.W. Harvey, K.C. Rush, A. Cockburn. John Wiley &
Sons Ltd.: Chichester, UK. 1999.
Generations
at Risk:
Reproductive Health and the Environment. T. Schettler, G. Solomon,
M. Valenti, A. Huddle. The MIT Press: Cambridge, MA. 1999.
Hormonal
Chaos: The
Scientific and Social Origins of the Environmental Endocrine Hypothesis.
S. Krimsky. Johns Hopkins University Press: Baltimore, MD. 2000.
| Endocrine
Disrupters - Journal Abstracts |
| National
Library of Medicine |
- Current approaches toward chemical mixture studies at the
National Institute of Environmental Health Sciences and the U.S.
National Toxicology Program. (1998)
- Sexual differentiation and environmental endocrine
disrupters. (1998)
- Health risk assessment of drinking water contaminants in
Canada:
the applicability of mixture risk assessment methods. (1997)
- Environmental estrogens and reproductive health: a
discussion of
the human and environmental data. (1997)
- The workshop on endocrine disrupter research needs: a
report. (1997)
- Relevance of risk assessment to exposed communities. (1995)
TITLE: Current approaches toward chemical mixture
studies at the National Institute of Environmental Health Sciences and
the U.S. National Toxicology Program.
AUTHORS: Bucher JR; Lucier G
SOURCE: Environ Health Perspect 1998
Dec;106 Suppl 6:1295-8
ABSTRACT: The National Institute of Environmental Health Sciences
(NIEHS) has several new initiatives involving chemical mixtures and has
recognized the need to develop new experimental approaches to enhance
our efforts in this area. Responding to recent increases in
nominations
of complex occupational exposures for toxicologic assessment by the
U.S. National Toxicology Program, the NIEHS and the National Institute
for Occupational Safety and Health have begun a program to characterize
exposures through field studies, identify biomarkers of exposure in
workers, and recreate relevant mixed exposures in
a laboratory setting. A second initiative with the National
Center for
Environmental
Health/Centers for Disease Control and Prevention will examine blood
samples
from the U.S. National Health and Nutrition Examination Survey
population
surveys for selected endocrine-disrupting agents and for common
patterns
of persistent xenobiotics, providing critical information for the
design
of animal studies to assess risks of relevant chemical mixtures to
humans.
New toxicology testing methods (lower cost, faster) will enhance our
ability
to study chemical mixtures (e.g., dioxin and dioxinlike chemicals,
combination
AIDS therapies). Ongoing method development efforts involve in vitro
functional
toxicology assays, screens for estrogenic activity, and carcinogenesis
studies
in transgenic mice. A major scientific initiative with mixtures
involves
studies of individual and mixtures of dioxin and dioxinlike chemicals
to
determine if toxic equivalence factors predict carcinogenic potency in
traditional
and transgenic bioassays. Complementing these studies is an
increased
emphasis
on physiologically based pharmacokinetic modeling, an activity central
to
the proper interpretation of chemical mixture studies.
TITLE: Sexual differentiation and environmental
endocrine disrupters.
AUTHORS: Toppari J; Skakkebaek NE
SOURCE: Baillieres Clin Endocrinol
Metab 1998 Apr;12(1):143-56
ABSTRACT: Male sexual differentiation is dependent on normal testicular
function, including secretion of testosterone from the Leydig cells,
and mullerian-inhibiting
substance from the Sertoli cells. External factors, such as
anti-androgens
and oestrogens, that disturb endocrine balance cause demasculinizing
and
feminizing effects in the developing male fetus. Oestrogens also
causes
adverse
effects in female fetuses, whereas anti-androgens have little
influence.
A growing number of chemicals have been found to possess either weak
oestrogenic,
anti- androgenic or other hormonal activities, and these are often
referred
to as
|
Note that
many of the
considerations we mentioned in discussing reproductive toxicity again
arise when examining the overall endocrine axes: lipid uptake by
steroidogenic tissues, actions of steroidogenic P450 enzymes
on
xenobiotics to produce proximal toxicants, interferences with
enterohepatic breakdown of lipoid hormones causing disruptions of
feedback loops, and modification of the molecular loads carried by
hormone binding globulins in circulation.
There are few
big surprises
here, mainly logical outcomes of the physical chemical and
physiological characteristics that make up the endocrine system itself.
We have
already covered the
physiology of the reproductive systems of the male and female including
the multiple levels of intercellular communication necessary for normal
function to occur. Note that this provides many targets for
potential
disruption by pharmacological or environmental agents. Regulation
by
the neuroendocrine components of the hypothalamus provide a target for
a variety of neurotoxic agents that may ultimately be expressed as
reproductive disruption and, therefore, toxicity. Secondary controls on
hypothalamic control such as the stress axis (adrenal axis) and leptin
regulation provide additional areas that may be disrupted by external
agents yet express themselves as reproductive problems.
Disruption of
the feedback regulation exerted by gonadal steroids on the hypothalamus
and hypophysis provide yet another target for disruption of
reproduction at
the level of the brain and pituitary. But potential targets
also exist
with
respect to the binding and actions of the gonadotropins at their
primary target
cells in the gonads, the cells supporting gamete production.
Steroid
disruptors: estrogen, androgen, progestin, corticoid
Environmental
Estrogens and Other
Hormones
Leydig cells
in the testis and
granulosa, thecal, stromal, and luteal cells in the ovary are all
steroidogenic. They make use of cholesterol and cholesterol ester
stores sequestered as lipid droplets to generate the gonadal steroids
that both support gonadal and reproductive tract function and provide
the systemic feedback to regulate hypothalamic and hypophysial control
of reproduction. Thus, these cells attract and concentrate
lipophilic
compounds that are capable of physically partitioning into fat. Brain
tissue also tends to be vulnerable in this way because of the
concentration of lipids in myelin. Oocytes themselves tend to
accumulate such materials because of their yolk content. Sperm
are
spared due to their comparatively low lipid content.
The Sertoli
cells and male
gametes are vulnerable to other agents. Those capable of
disrupting
tight junction formation may break down the junctional complexes
between Sertoli cells
and developing spermatogenic cells, disrupting the blood testis barrier
that is needed to maintain integrity of the semeniferous tubule
compartments and keep spermatogenesis well orchestrated. In
contrast,
the granulosa cells (especially of the cumulus layer) and the oocyte
are vulnerable to agents that disrupt gap junctional complexes since
these are needed to coordinate the functions of the ovarian follicle
and the growth of the oocyte.
While the
spermatogenic cells
are particularly sensitive to toxins affecting rapidly growing cells,
mammalian oocytes are sensitive to agents that interfere with both
meiosis and apoptosis (the normal manner in which oocytes begin their
decline toward atresia). Agents
that speed the loss of oocytes can lead to premature depletion of the
oocyte
pool because that pool is physiologically limited in size at mammalian
birth.
Spermatogenesis on the other hand seems relatively robust but can still
be
disrupted by any agent capable of modifying the sequence of events
leading
to formation of normal spermatozoa or their transport to the female
tract.
The female
tract itself is a
sensitive target because of the cyclical proliferation,
differentiation, and regression that takes place in the tissues of the
endometrium, and, to a lesser extent, in the vagina. Steroid
mimics,
agents that disrupt cellular proliferation (or regression), or those
that alter paracrine signal transmission among the
tissues of the female tract can change the timing or movement of
gametes and/or
zygotes and blastulae through the female system. Changes in
ciliary
beat
in the Fallopian tube, changes in fimbrial activity, alterations in
cervical
mucus secretion or biochemistry, alterations of female tract
contractility or contractile responses to seminal prostaglandins can
all affect the success of gamete movement, fertilization, and/or
implantation and gestation.
Finally, any
agents that
disrupt enterohepatic
metabolism of steroids or other reproductive tract signaling agents
will
have potential reproductive toxicity effects. Either they will
cause
steroids
to be cleared too rapidly and therefore will alter the normal
conversation
between the gonads and the hypothalamus/pituitary or they will cause
steroids
to be cleared too slowly, similarly disrupting this set of feedback
equilibria.
Such impacts would include alterations of hepatic production of serum
binding
globulins. Sex-hormone binding globulin/testosterone-estradiol binding
globulin
(SHBG/TeBG), corticoid binding globulin (CBG), thyroid-binding globulin
(TBG),
and thyroid-binding prealbumin/transthyretin (a binder for both TBG and
retinoic
acid derivatives) levels all respond to circulating hormone
levels.
SHBG,
CBG, and TBG all increase in response to bioavailable estradiol levels;
they
decrease in response to androgens (or to the decline in estradiol to
androgen
ratio). SHBG and TBG also seem to increase with circulating
thyroid
hormone
levels. Finally, CBG binds both progesterone and corticoids
efficiently
while
it SHBG binds preferentially to androgens. Thus, endocrine
steroid
mimics
not only change the dynamics of steroid binding to receptors in target
cells,
they also may alter bioavailable endogenous hormone levels by changing
serum
buffering capacity for these hormones and/or competing for that
capacity.
In testing
for impacts on the
endocrine system we again run into the problem seen with genetic and
developmental
toxicity. To determine if the toxin is effecting the parents,
offspring
must
be generated. But because the offspring may themselves be
carriers of
the
toxic defect imposed by exposure of the parents to the toxic agent, the
offspring
must be evaluated as well as the parents for anomalies of structure and
function. And function includes reproductive function. Thus,
reproductive toxicity tests,
as they exist at the present time, include evaluation through three
generations
(parental, F1, and F2) to ascertain what the
impacts on reproduction
might
be.
A couple of
additional articles
also help to demonstrate the scope of a small part of the endocrine
disruption literature, these are linked to the posted session.
[An article
concerning
disruption of sexual development in birds on the West coast as a result
of exposure to
chlorinated hydrocarbons.]
[An article
from the Ecologist
regarding possible lindane contamination of chocolate from Ghana as a
result of economically driven agricultural practices.]
These two
articles remind us
that the legacy of synthetic polychlorinated hydrocarbons is still with
us. Even 40 years after the publication of Silent Spring
and
the governmental actions in setting up the EPA, NIEHS, and
strengthening the FDA, these pesticides and their relatives remain a
persistent problem. The impact of chlorinated aromatics including
DDT
and PCBs has continued to unfold since their development in the 1930s
and their association with the decline of predatory bird species
described in the 1960s and early 1970s. Dr. Jeremy Hatch of
UMB/Biology
works on a population of roseate terns who have a decreased fertility
and marked decrease in the proportion of males due to contamination of
food sources near
Cape Cod with chlorinated hydrocarbons. The Wisconsin Raptor
Center and
the
WARF Institute played a major role in describing the decline of
predatory bird species as a result of egg-shell thinning, calcium
metabolism problems, and vitality reduction associated with
accumulation of excessive levels of these compounds in bird fat and egg
yolk. The compounds accumulate up the food chain, concentrate in
fatty
tissues, inhibit the carbonic anhydrase necessary
for egg shell formation, and tend to be "off-loaded" in the fatty yolk
of
the eggs. Thus, young are exposed to even higher levels than are adults
during
crucial times of development. Although PCBs were in use when DDT
was
being
identified with bird population problems, it wasn't until the mid- to
late
1970s when it became fully apparent that this series of widely used
electrically
nonconductive, fire retardant, heat dispersing agents was leaking into
the
environment in significant quantities and generating many of the same
kinds
of problems the chlorinated hydrocarbon pesticides had.
The efficacy
of DDT as an
insecticide was legend in the period following World War II. But
by the
late 1960s most areas using it to suppress malaria bearing mosquitoes
noted a marked decline in its impact due to development of
DDT-resistant strains of Anopheles and Aedes species. Interestingly, a
report on 5/4/03 on National Public Radio described the
recent, apparently successful, house-spraying program now being used in
South Africa to decrease malarial infections. Because this
program is
seeing success it is being extended. One must only wonder how
long it can remain successful and how severe an impact it will have on
native non-target species as well as polar species where these
compounds tend to ultimately accumulate.
All of these
compounds are
resistant to degradation but gradually give way to the actions of mixed
function oxidases and/or exposure to intense UV irradiation or to high
heat. They are slowly oxidized to related ketones, carboxylic
acids,
and alcohols that are themselves slowly broken down by P450
type
enzymes and/or conjugated to glucuronic acid, sulfate, or amino acids
to yield more water soluble compounds. Along the way,
however, these compounds and some of their metabolites may mimic
estrogens or thyroid hormones.
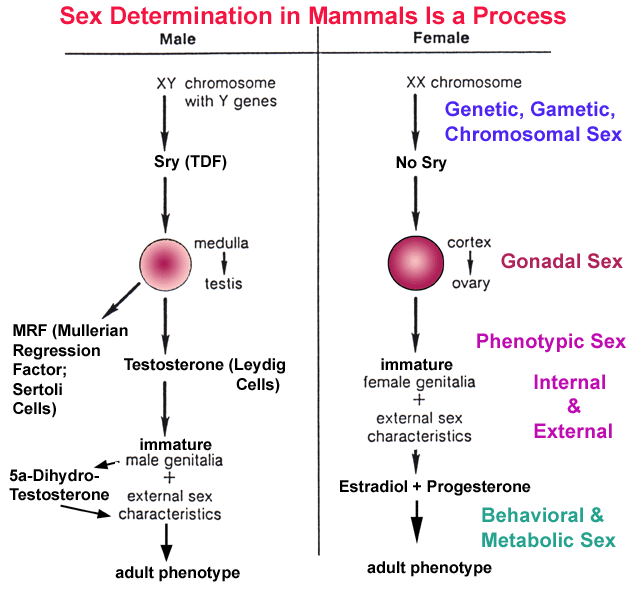
Estrogen:
The classic
example of a
synthetic entering the environment and food chain and having a marked
toxicological
 impact is
provided
by the actions of diethylstilbestrol. This compound is a potent
synthetic
estrogen that acts via the same
mechanisms as estradiol (but is not
buffered
by binding to the sex hormone binding globulin that modulates free
levels
of estradiol). It binds to intracellular estrogen receptors with
an
affinity
approximately 100 times higher tha n
the endogenous ligand 17-estradiol.
When
given during early to mid-gestation to prevent early miscarriage this
compound
proved a developmental toxicant. Female children of treated women
demonstrated
a range of reproductive tract anomalies up to and including altered
cervical
cell lineages that eventually gave rise to cervical cancers. Male
children
also demonstrated impacts ranging up to hypospadiasis (incomplete
ventral
fusion of the tissue folds forming the penis). Given the
multistage
nature
of the sexual differentiation process [genetic sex -- being
positive
for SRY > gonadal sex -- gonadal morphogenesis from the
indifferent
gonad medulla in males or cortex in females > differentia
impact is
provided
by the actions of diethylstilbestrol. This compound is a potent
synthetic
estrogen that acts via the same
mechanisms as estradiol (but is not
buffered
by binding to the sex hormone binding globulin that modulates free
levels
of estradiol). It binds to intracellular estrogen receptors with
an
affinity
approximately 100 times higher tha n
the endogenous ligand 17-estradiol.
When
given during early to mid-gestation to prevent early miscarriage this
compound
proved a developmental toxicant. Female children of treated women
demonstrated
a range of reproductive tract anomalies up to and including altered
cervical
cell lineages that eventually gave rise to cervical cancers. Male
children
also demonstrated impacts ranging up to hypospadiasis (incomplete
ventral
fusion of the tissue folds forming the penis). Given the
multistage
nature
of the sexual differentiation process [genetic sex -- being
positive
for SRY > gonadal sex -- gonadal morphogenesis from the
indifferent
gonad medulla in males or cortex in females > differentia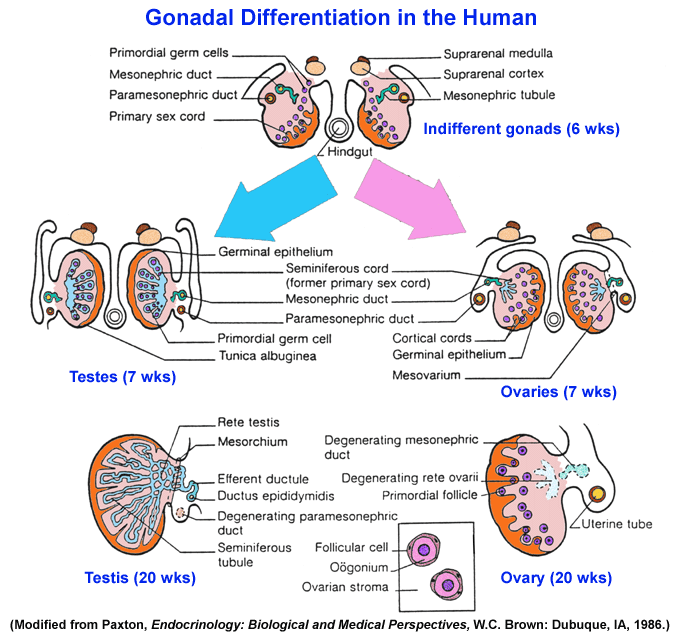 tion in the early testis of Sertoli
cells
that produce Anti-Mullerian Hormone
and Leydig cells that produce
testosterone
>
internal phenotypic sex --
suppression of Mullerian ductal
derivatives
including
Fallopian
tubes and uterus in the 1st trimester male fetus combined
with
the androgenic support of the further differentiation of the Wolffian
ductal
derivatives to produce the interior components of the male reproductive
tract
including the epididymis, vas deferens, and seminal vesicles & external
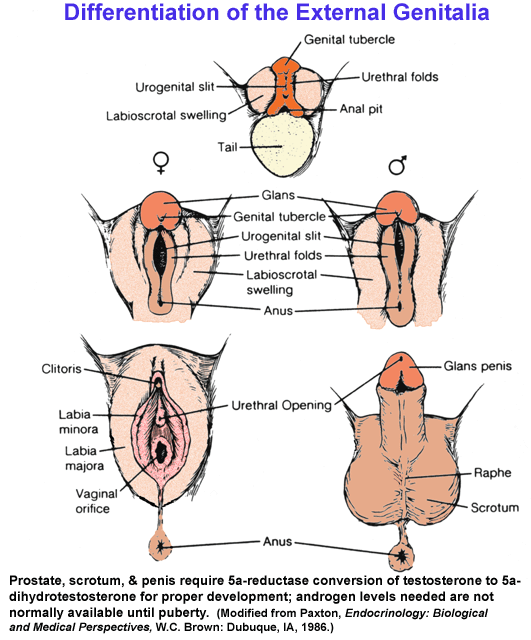
phenotypic sex - prenatal testosterone and 5-dihydrotestosterone,
generated
within target tissues, masculinize external genitalia, causing
formation
of the prostate, penis, and scrotum, as well as internal organs such as
liver
and kidney, while peripubertal androgens and estrogens drive
morphological
maturation > brain sex - appears to be largely defined by
estrogen
generated in the target cells during the natal/perinatal period in both
females
and males while later behaviors seem to respond preferentially to
estrogens
in females and androgens in males in mammals it
is perhaps only surprising that more
marked impacts failed to occur
tion in the early testis of Sertoli
cells
that produce Anti-Mullerian Hormone
and Leydig cells that produce
testosterone
>
internal phenotypic sex --
suppression of Mullerian ductal
derivatives
including
Fallopian
tubes and uterus in the 1st trimester male fetus combined
with
the androgenic support of the further differentiation of the Wolffian
ductal
derivatives to produce the interior components of the male reproductive
tract
including the epididymis, vas deferens, and seminal vesicles & external
phenotypic sex - prenatal testosterone and 5-dihydrotestosterone,
generated
within target tissues, masculinize external genitalia, causing
formation
of the prostate, penis, and scrotum, as well as internal organs such as
liver
and kidney, while peripubertal androgens and estrogens drive
morphological
maturation > brain sex - appears to be largely defined by
estrogen
generated in the target cells during the natal/perinatal period in both
females
and males while later behaviors seem to respond preferentially to
estrogens
in females and androgens in males in mammals it
is perhaps only surprising that more
marked impacts failed to occur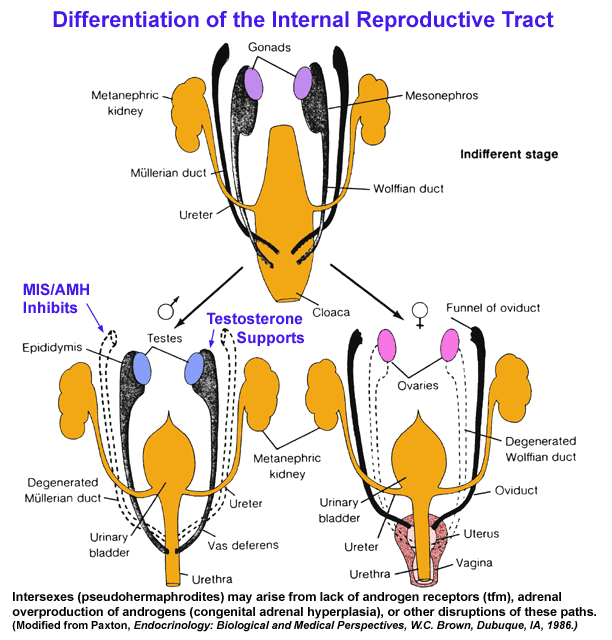 in male offspring.
in male offspring.
And while pharmacological
or environmental
exposures
to synthetic estrogens are described
above, a variety of
reports now discuss phytoestrogens as one natural source of ecological
exposure that may help explain different reproductive and breast cancer
rates across animal populations including humans. Soy extracts
are
being sold as a natural alternative source of estrogenic substances
that can help alleviate the undesirable symptoms associated with
menopause, e.g., hot flashes,
drying and thinning of skin and vaginal
linings. Meanwhile a variety of plant oils and extracts are being
used
in skin lotions to combat the hormonal impacts of aging.
Breast tissue, specifically the
lactogenic alveolar tree, is a prime target for estrogen actions in
the adult. Since this area of the
nulligravid
adult female breast is prepared to respond to estrogen by
extensive proliferation in
preparation
for lactation, it is perhaps not surprising that nulligravid women are prarticularly
prone to development of
breast
cancers in their mid- to late reproductive years. Early childbeari
 ng
has
repeatedly been associated
with decreased risk for breast cancer. One idea that has now been
tested
was that the cumulative impact of
repeated
exposure of breast, cervical, and endometrial tissues to estradiol
without
a prolonged period of intervening elevated progesterone (as occurs in
pregnancy)
was capable of triggering enough cellular proliation
to induce neoplastic tissue
growth.
By suppressing these actions with an anti-estrogen, tamoxifen, given
prophylactically,
the hope was that rates for these cancers would drop in the test
population.
While the inherent estrogenicity of the tamoxifen or the impact of its
interferences
with estradiol, elevated the rates of some reproductive tract cancers
in
a broadly based study, this was not evident in a study of women at high
risk
for breast cancer due to family genetic background and/or life history,
including
early menarche and nulliparity. Part of the prophylactic effects
of
phytoestrogens,
and a possible hormetic impact of weak synthetic estrogens may be due
to
suppressive impacts similar to tamoxifen.
ng
has
repeatedly been associated
with decreased risk for breast cancer. One idea that has now been
tested
was that the cumulative impact of
repeated
exposure of breast, cervical, and endometrial tissues to estradiol
without
a prolonged period of intervening elevated progesterone (as occurs in
pregnancy)
was capable of triggering enough cellular proliation
to induce neoplastic tissue
growth.
By suppressing these actions with an anti-estrogen, tamoxifen, given
prophylactically,
the hope was that rates for these cancers would drop in the test
population.
While the inherent estrogenicity of the tamoxifen or the impact of its
interferences
with estradiol, elevated the rates of some reproductive tract cancers
in
a broadly based study, this was not evident in a study of women at high
risk
for breast cancer due to family genetic background and/or life history,
including
early menarche and nulliparity. Part of the prophylactic effects
of
phytoestrogens,
and a possible hormetic impact of weak synthetic estrogens may be due
to
suppressive impacts similar to tamoxifen.
Androgen:
Note that
estrogenic impacts
also include a wide range of alterations of metabolic parameters
including circulating lipid profiles (decrease LDL) and bone calcium
deposition (suppressed by estrogens). Since androgen frequently
antagonizes these changes it is often difficult to distinguish in whole
animal models between the anti-estrogenic effect of a compound and its
androgenic effects. Likewise, it is similarly difficult to
distinguish
between an anti-androgenic effect and an estrogenic effect. It is
largely due to this ambiguity that there is a much more limited
literature dealing with androgenic or anti-androgenic effects of
xenobiotics. This is also compounded by the fact that several
different
androgenic steroids can bind to intracellular androgen receptors with
differing, but often sufficient, affinity to induce transcriptional and
translational changes, testosterone, 5-dihydrotestosterone, and
androst-4,5-ene-3,17-dione can all interact with the androgen receptor.
The
calculated impacts of
cumulative exposures to environmental estrogens cannot explain some of
the information marshaled as evidence for endocrine disruption in
males. The downward secular trend in human male sperm counts has
been
suggested as one such index (though the possibility that better
techniques have provided better, but lower counts cannot be totally
discarded even now). Another is the inability of endogenous
androgens
to block nipple development or to support internal and external sex
phenotypic development in rats (as it normally does) in animals dosed
with suspect compounds. On the basis of this evidence, p,p'-DDE,
metabolites of vinclozolin, and di-n-butyl phthalate have all been
designated anti-androgens.
Progestins
& Corticoids:
Note that
progestins and
glucocorticoids share rather similar chemical structures and solubility
properties. Their receptor proteins (progesterone receptor, PR,
and
glucocorticoid receptor, GR) are likewise quite similar; both sets of
compounds are capable of binding, with lower affinity than the major
ligand, to each other's receptors. Moreover, that binding can
trigger
the complementary biological activity. Thus, for example, given
high
enough concentrations of progesterone, immune suppression can be
induced via activation of the glucocorticoid receptors. In addition,
both steroids can bind to the mineralocorticoid receptor (MR) that
normally binds aldosterone. The usual physiological barrier to
this
happening in all MR-containing tissues is the presence of the enzyme
11-hydroxysteroid dehydrogenase that metabolizes cortisol to
cortisone.
Finally, both glucocorticoids and progestins bind rather effectively to
CBG and, in those species such as guinea pig that have it, to PBG,
progesterone binding globulin. So they can displace or replace
one
another in a number of biological situations.
One class of
pharmaceuticals is
know to act as anti-progestins/anti-glucocorticoids, e.g., RU486,
mifepristone. Another is used in oral
contraception/postmenopausal
hormonal replacement to replace intrinsically generated progesterone
(medroxyprogesterone, norethindrone, norgestrel, norethynodrel,
norgestimate) and seems to counteract the positive effects that
estrogen has on cardiovascular parameters. A third class
comprises
synthetic glucocorticoids: prednisone, prednisolone, dexamethasone,
triamcinolone) that are immunosuppressive and promote cardiac
hypertrophy and fibrosis at toxic levels. Most of the clinical
side-effects among the agonistic drugs derive from occupation of
non-targeted receptors, PR and MR for glucocorticoids, GR and PR for
progestins. Their impacts on organisms exposed to high levels of
the
drugs in effluents or untreated sewage streams derive from both their
targeted and their untargeted agonistic actions. The antagonist
functions by blocking both PR and GR. Fortunately most of these
compounds contain enough functional groupings to allow them to be
catabolized and excreted rather efficiently.
Arsenic has
also been described
as an endocrine disruptor.
The issue of arsenic as an endocrine
disrupting agent
is somewhat puzzling. Still, arsenic's position in the periodic
table
suggests
it may interact and/or interfere with metabolic processes involving
phosphorylation.
As this is common during many transduction processes including the
movement
of glucocorticoid receptors to the nucleus with the concomitant
dissociation
of a series of heat shock proteins, it may not be so surprising that
arsenic
may have some specificity in its actions in such systems. It may
also
be
possible that arsenate's resemblance to vanadate plays a role because
vanadate
is commonly used during in vitro experimentation to stabilize isolated
glucocorticoid
receptors. Such stabilization reduces hormone-receptor turnover
and may
lock
the expression of certain genes either "on" or "off."
Thyroid,
retinoid and other disruptors
Thyroid:
Exposures to
PCBs (and the
structurally related polybrominated biphenyls, PBBs) can potentially
alter thyroid function in part because of the structural similarities
of some of the PCB isomers and thyroxine or triiodothyronine.
About 50%
of thyroxine is carried in circulation by thyroid binding globulin,
TBG, while 45+% is carried by albumin or
transthyretin. Much
of thyroxine enters cells by diffusion, but a substantial quantity also
enters via the aromatic amino acid transport proteins in cell
membranes. After cell entry, thyroxine is de-iodinated to
triiodothyronine which then binds to receptors that usually reside on
DNA binding sites within the nucleus; binding often results in release
of the hormone receptor complex from DNA and in a change in DNA
structure and transcriptional activity. What are the obvious
possible
targets for PCB toxic effects? If we include molecular clearance
pathways, are there other targets? What if the toxicant is a
slightly
acidic derivative of a PCB? What about an amine derivative of a
PCB?
Note the
molecules needed to
synthesize thyroxine, transport it to its somatic targets, activate it
in target cells by removing an iodine atom, allow it to activate genes
via binding to a receptor sitting on a thyroid recognition element
within the DNA, and terminate its actions via oxidation, conjugation
and an increased water solubility and urinary
elimination. If we see a structural similarity of a compound to a
known
hormone
or physiological compound, the obvious place to look for potential
molecular
targets are those molecules normally involved in that compound's
physiology
and actions. In the absence of a structural similarity, but with
a
known
set of end effects, the targets may lie on the paths leading to normal
activations
of the ultimately affected tissue. As this often involves
hormones,
hormonal
pathways may well be fruitfully explored.
Retinoid
& Other:
Although most
attention has
been directed
to the issue of environmental estrogens and/or environmental
anti-androgens, thought should also be given to compounds that have
potential to disrupt the
thyroid axis, vitamin D metabolism, retinoid growth regulation, and the
adrenal
axis.
Coverage in
C&D pays
particular attention to tumorigenesis and induction of neoplasias in
the endocrine tissues. It also notes, however, that many of these
effects are the indirect result of disruptions of normal homeostatic
feedback loops with much of the hyperplastic induction being due to
chronic elevations in trophic hormone production and target tissue
stimulation. While impacts that produce neoplasias are definitely
dramatic, they are really the extreme manifestations of functionally
more important disruptions of endocrine homeostasis. Considerable
note
was taken of the apparently higher susceptibility of rodents,
particularly some of the
favored toxicity testing strains such as Fischer 344 rats, to formation
of
endocrine tumors relative to what is seen in other rodent strains, or
other
species including humans. Is it worth considering how functional
tests
of
the endocrine axis might serve as more sensitive biomarkers for toxic
insults
to these systems than waiting for rats to develop visible tumors?
Further,
although healthy adult
reproductive function is required for species health, it is not a
requirement of individual health. On the other hand, pancreatic,
parathyroid, and kidney/adrenal glomerular function are required for
individual well-being. In the absence of the internal homeostasis
governed by these systems no individual would be capable of surviving
to reproduction much less successfully reproducing. Shouldn't
more
emphasis be placed on these systems vis-a-vis
endocrine disruption if
the promotion of health of the population are among our goals?
Take
Home Exam Questions (Due Next Week)
© 2005
Kenneth L. Campbell